Family Davidiaceae
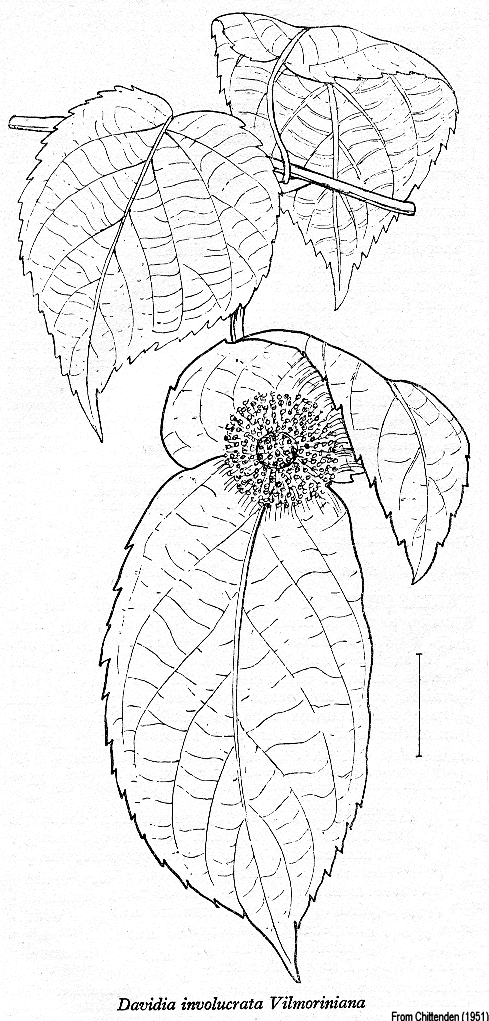
Davidiaceae Takht.~ Cornaceae, Nyssaceae Habit and leaf form. Trees. Leaves deciduous; alternate; simple. Lamina entire; broadly ovate; pinnately veined; cross-venulate; cordate. Leaves exstipulate. Lamina margins dentate. Leaf anatomy. Lamina dorsiventral. Minor leaf veins without phloem transfer cells. Stem anatomy. Cork cambium present; initially superficial. Internal phloem absent. Secondary thickening developing from a conventional cambial ring. ‘Included’ phloem absent. Vessel end-walls scalariform. Wood parenchyma apotracheal. Reproductive type, pollination. Plants andromonoecious. Female flowers with staminodes (‘the female-fertile flowers with few to many small staminodes inserted halway up the ovary’ (Airy Shaw 1973)). Gynoecium of male flowers absent. Inflorescence, floral, fruit and seed morphology. Flowers aggregated in ‘inflorescences’; in heads. Inflorescences terminal; dense, terminal, pedunculate, globular capitula, each with many scarcely distinct male flowers and a single, obliquely terminal hermaphrodite (or female?) one; with involucral bracts (each capitulum subtended by a pair of large, unequal, drooping, white, petaloid bracts); pseudanthial. Perianth vestigial (perhaps, in the hermaphrodite flowers), or absent (the male flowers consisting of stamens only, the female more or less limited to an ovary and stamens (plus staminodes?)). Androecium in male flowers, 5–6. Androecial members free of one another. Androecium exclusively of fertile stamens (in male flowers), or including staminodes (in the terminal, hermaphrodite flower, according to Airy Shaw). Stamens 5–6; filantherous (the filaments long). Anthers dehiscing via longitudinal slits; tetrasporangiate. Pollen grains aperturate; 3 aperturate; colporate. Gynoecium 6–10 carpelled. The pistil 6–9 celled. Gynoecium syncarpous; synovarious to synstylovarious; inferior. Ovary 6–9 locular. Epigynous disk absent. Gynoecium stylate. Styles 1 (columnar, exceeding the androecium). Stigmas 6–10 (‘the style with 6–10 branches’); dry type; non-papillate; Group II type. Placentation axile. Ovules 1 per locule; pendulous; epitropous; with ventral raphe; anatropous; unitegmic; more or less crassinucellate. Endothelium differentiated. Endosperm formation cellular. Fruit fleshy; indehiscent; a drupe (pear-shaped, about 3.5 cm long, with granular mesocarp). The drupes with one stone (endocarp bony, longitudinally sulcate, 3–5 locular). Seeds endospermic. Endosperm oily. Embryo well differentiated (rather large). Cotyledons 2. Embryo straight. Seedling. Germination phanerocotylar. Physiology, biochemistry. Not cyanogenic. Iridoids detected; ‘Route I’ type (+seco). Verbascosides not detected. Proanthocyanidins absent. Flavonols present; kaempferol and quercetin. Ellagic acid present. Arbutin absent. Saponins/sapogenins present and absent. Geography, cytology. Holarctic. Temperate. China. N = 21. Taxonomy. Subclass Dicotyledonae; Tenuinucelli (?). Dahlgren’s Superorder Corniflorae; Cornales. Cronquist’s Subclass Rosidae; Cornales. APG 3 core angiosperms; core eudicot; Superorder Asteranae; Order Cornales (as a synonym of Cornaceae). Species 1. Genera 1; only genus, Davidia. See Eyde (1988). This family exemplifies the well known difficulties in distributing certain Dicot families between Dahlgren’s Araliiflorae and Corniflorae. It is equally hard to assign them with confidence to the higher level groupings Crassinucelli and Tenuinucelli. This is interesting, given that the latter evidently represent a major divergence in the Dicot line of descent (cf.Young and Watson 1970, Chase et al. 1993). Illustrations. |
Support our developers





{kind=link}